
第一作者:林滨城广东工业大学生态环境与资源学院
通讯作者:马金星教授广东工业大学生态环境与资源学院
第一单位:广东工业大学生态环境与资源学院
论文DOI:10.1016/j.watres.2025.123326
图文摘要
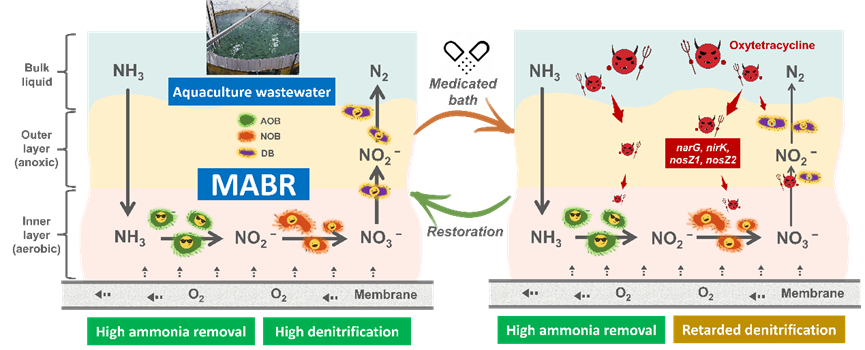
论文简介
近日,我院马金星教授课题组在环境领域知名学术期刊Water Research上发表了题为“Stratified biofilm structure of MABR enabling efficient ammonia removal from aquaculture medicated bath wastewater”的研究论文。
水产养殖过程需要使用抗生素来治疗或预防寄生虫和细菌感染。治疗方式主要包括将抗生素作为辅助剂加入饲料中或使用药浴。当养殖生物表现出与疾病相关的厌食症状时,药浴可能是唯一有效的治疗手段。然而,水产养殖药浴废水中残留的高浓度抗生素对传统生物脱氮工艺提出了挑战。本研究以低能耗、具有生物分层结构特征的膜曝气生物膜反应器(MABR)为研究对象,考察多生态位功能菌群协同处理水产养殖药浴废水的效果。实验结果表明,即使在高浓度土霉素(OTC)胁迫条件下,MABR仍可实现98.2±1.8%的氨氮(NH4+-N)高效去除,该性能归因于生物膜对内层功能菌群(如氨氧化菌AOB与亚硝酸盐氧化菌NOB)的保护作用。硝化过程相关的基因(amoA/B和nxrA)表达整体呈上调趋势,外排泵激活机制协同形成微生物应激响应。然而,尽管Denitratisoma oestradiolicum(14.5%)和反硝化细菌克隆NOA-1-C(41.7%)显著富集,但总氮(TN)去除效率从95.3±2.5%降至76.0±8.8%,这可能是由于narG基因表达受限所致。终止OTC投加并调整运行参数后,总氮去除率恢复至87.4±5.8%。本研究结果展示了MABR技术在水产养殖药浴废水处理中的有效性和适应性。
图文导读
经170天长期运行后,MABR系统展现出优异的污染物去除性能(图1a-c)。在15 mg L⁻¹ OTC长期胁迫条件下,系统通过分层生物膜结构维持NH4+-N去除率达98.2±1.8%(图1a),而TN去除率从初始95.3%下降至76.0%(图1c),表明反硝化过程受到抑制。通过调控曝气压至7±1 kPa后,成功构建缺氧微环境(DO=0.6 mg L⁻¹,图1d),促使TN去除率恢复至87.4±5.8%。值得注意的是,主体溶液DO水平的降低未显著影响生物膜内层AOB活性,这突显了分层生物膜的独特优势,有力证实了MABR系统中同步硝化反硝化的有效性。
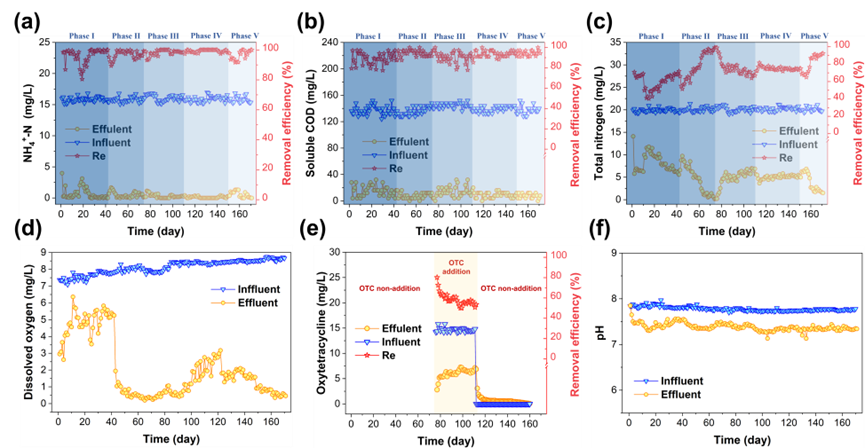
图1:MABR系统170天运行性能。(a)氨氮、(b)COD及(c)总氮浓度与去除率;(d)溶解氧(DO)、(e)土霉素浓度与去除效率及(f)进/出水pH值。注:进/出水浓度对应左轴,去除率(Re)对应右轴。
CLSM三维成像显示,OTC暴露15天后蛋白质、多糖及脂质产量较未暴露组显著增加(图2b3),表明微生物通过快速合成胞外聚合物(EPS)建立防御机制。伴随EPS的累积,生物膜厚度由初始1.14 mm增至1.70 mm(图2b1-2),验证了结构强化与抗逆性提升的关联性。持续暴露至胁迫后期,生物膜厚度趋于稳定(图2c1-2)且代谢活性维持动态平衡,说明微生物群落已形成新的稳态。值得注意的是,此时多糖含量显著降低而脂质占比升高(图2c3),这种EPS组分比例的重分配现象,可能反映了微生物从急性应激(依赖多糖类物质快速成膜)向慢性适应(增强脂质介导的膜稳定性)的代谢策略转变。
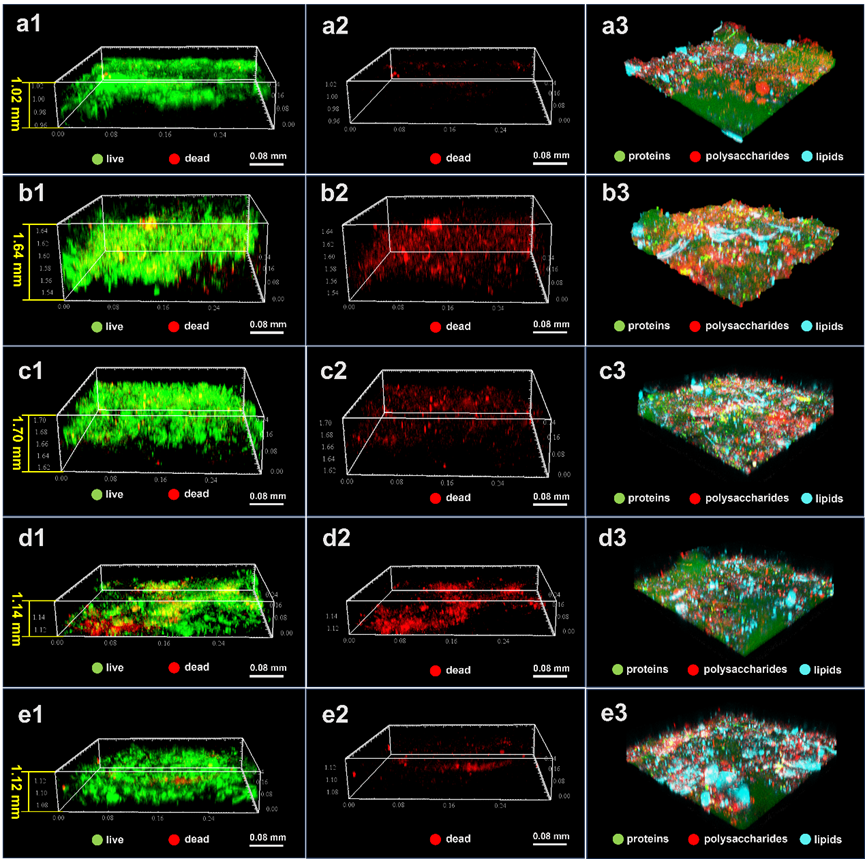
图2:MABR运行过程中生物膜生长与组分动态可视化。(a1-a3)成熟生物膜,(b1-b3)OTC暴露15天,(c1-c3)OTC暴露35天,(d1-d3)修复期及(e1-e3)气压调节后的生物膜活/死细胞活性染色及蛋白质、多糖、脂质分布。注:活/死细胞染色:a1-e1示活细胞(绿色荧光),a2-e2示死细胞(红色荧光)。组分染色:a3-e3中蛋白质、多糖、脂质分别以绿色、红色、靛蓝显色。
微生物多样性分析揭示OTC暴露35天的生物膜分区响应特征:内层菌群丰度波动幅度小于5%,而外层拟杆菌纲(Bacteroidia)相对丰度较S6阶段提升2.6倍,同时γ-变形菌纲(Gammaproteobacteria)丰度下降至原水平的48.8%。这种空间分异现象表明,拟杆菌纲可能通过外膜修饰等机制获得OTC抗性,而作为关键反硝化功能菌的γ-变形菌纲衰减,或直接导致系统TN去除率下降(图1c)。在物种水平上,内层样本S7中Denitratisoma oestradiolicum丰度增长2.83倍,而外层S8该菌种占比飙升至70.9%,暗示生物膜通过空间功能菌群重组维持脱氮能力。LEfSe分析进一步确认胁迫组功能菌的显著富集(LDA>3,p<0.01)。
通过高通量定量PCR(HT-qPCR)揭示生物膜内氮循环功能基因的分层表达特征。氨单加氧酶基因(amoA/B)在内层的拷贝数密度较外层高43%(图3a),这解释了系统在长期胁迫下仍维持98.2±1.8%的NH₄⁺-N去除率的现象(图1a)。亚硝酸盐氧化基因(nxrA)表达量显著增加,导致中间产物NO₂⁻-N被快速转化为NO₃⁻-N(检测浓度<0.5 mg L⁻¹)。短期胁迫下nirS1/2及nirK1-3拷贝数激增,支撑系统出水的低NO2−-N水平(图3c),厌氧氨氧化基因(hzsB)在内层富集,推测生物膜内局部微缺氧区形成,这促进了Anammox的性能发挥(图3d)。停止胁迫并调控曝气后,硝酸盐还原酶基因(narG)基因表达回升(图3b),驱动MABR反硝化性能恢复,印证功能基因动态对工艺调控的响应机制。
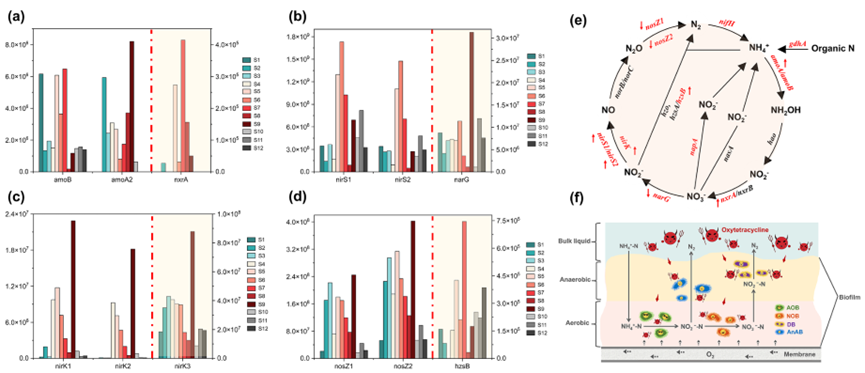
图3:基于高通量qPCR基因芯片测序的微生物群落中元素氮循环基因的演化。负责(a)氨和亚硝酸盐氮氧化,(b)硝酸盐氮和亚硝酸盐氮还原,(c)亚硝酸盐氮转化为一氧化氮,(d)反硝化过程中一氧化二氮转化的功能基因;(e)上调和下调N循环途径的示意图;(f)高OTC胁迫对MABR中N循环影响。注:S1:接种污泥;S2:成熟生物膜(阶段I);S3:阶段II内层生物膜;S4:阶段II外层生物膜;S5:OTC暴露15天(阶段III)内层;S6:OTC暴露15天外层;S7:OTC暴露35天(阶段III)内层;S8:OTC暴露35天外层;S9:修复期(阶段IV)内层;S10:修复期外层;S11:气压调节后(阶段V)内层;S12:气压调节后外层。在图e中,所有样品中注释的基因用红色表示,而未成功注释的基因用黑色表示;在图f中,AOB:氨氧化细菌,NOB:亚硝酸盐氧化细菌,DB:反硝化细菌,AnAB:厌氧氨氧化细菌。
KEGG代谢通路分析显示(图4),硝化作用相关代谢通路(如K10944–K10946和K10535)在抗生素胁迫下未呈现显著差异,证实高浓度抗生素对MABR内硝化过程的代谢通路水平影响微弱。与此相反,反硝化相关通路(K00370、K02567、K00368、K15864、K04561及K00376)在抗生素胁迫下发生显著改变,宏基因组分析结果与qPCR基因芯片定量数据具有良好一致性(图4)。
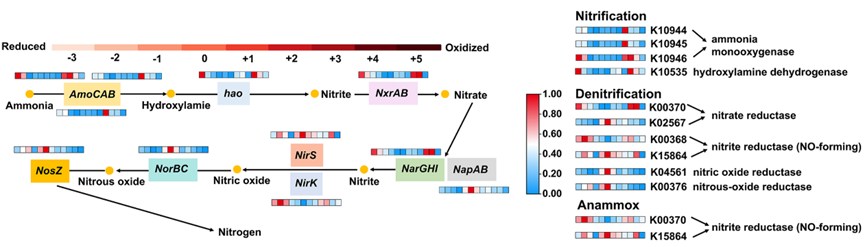
图4:使用宏基因组分析对MABR系统氮代谢途径在OTC应激下的KEGG直系同源性(KO)。垂直比例尺表示KEGG KO注释的标准化丰度。注:条形图(0.00–1.00)表示KEGG相关数字的相对丰度。蓝色色调表示相对丰度较低,而红色色调表示相对丰度较高。
基于CARD数据库的注释分析揭示,OTC胁迫诱导抗生素抗性基因(ARGs)发生显著空间重组(图5)。β-内酰胺酶类抗性基因在内外层呈现差异化表达,可能与生物膜内菌群间抗性基因水平转移(HGT)引发的共抗性效应相关(图5a)。OTC暴露初期(S5-S6阶段),外层菌群因直接接触高浓度OTC(15 mg L⁻¹),其外排泵基因丰度显著高于内层(图5b),表明空间暴露梯度驱动主动外排防御策略。持续胁迫至S7-S8阶段,外层菌群外排泵丰度出现下降且低于内层,推测其防御能力已达外排防御阈值。停用OTC后(S9-S10),外排泵基因丰度整体下降,经曝气压优化(S11-S12阶段)最终恢复至胁迫前水平。多元抗性机制分析表明(图5c),系统通过三重防御机制缓解OTC毒性:①靶点修饰;②酶解失活;③膜通透性调控协同降低抗生素摄入。
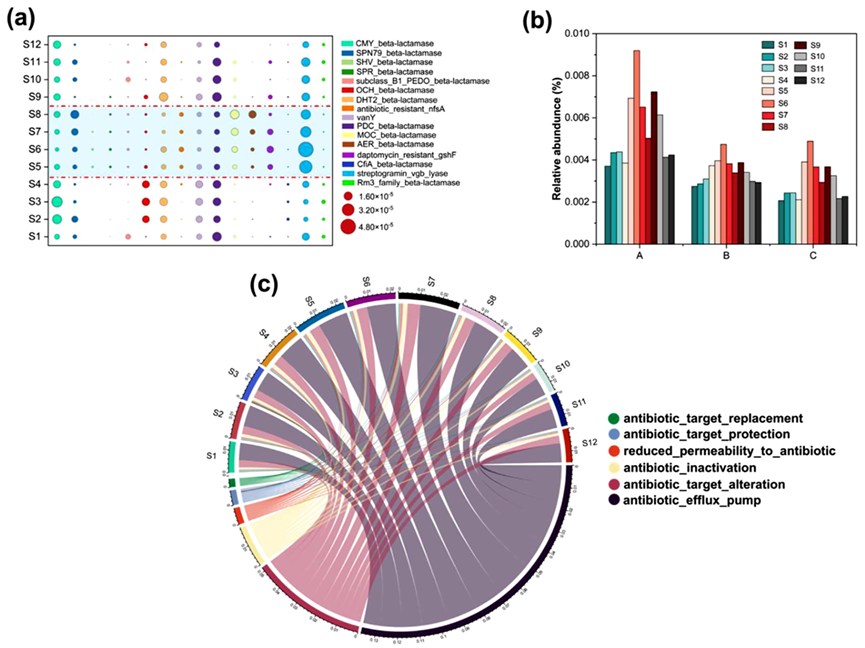
图5:抗性基因的演化。(a)不同类别和(b)外排泵基因的相对丰度;(c)基于综合抗生素抗性数据库(CARD)的不同时期抗性机制弦图。注:S1-S12与图3中描述的一致;在图b中,A:抗性结瘤细胞分裂(RND)抗生素外排泵,B:ATP结合盒(ABC)抗生素外排泵,C:主要促进剂超家族(MFS)抗生素外排泵。
小结与展望
本研究通过评估MABR长期运行中分层生物膜结构对水产养殖药浴废水的脱氮效果。实验表明:高浓度OTC暴露可导致生物膜活性降低及EPS组分显著重构。硝化过程受抑效应微弱(NH4+-N去除率持续>93.2%),而反硝化性能显著受损(TN平均去除率降至76.0±8.8%),其机制主要源于生物膜外层反硝化功能基因(narG、nirK、nosZ1、nosZ2)拷贝数整体下调。该代谢异质性可解释反硝化菌(如Denitratisoma oestradiolicum、反硝化富集菌NOA-1-C)丰度增加与脱氮效率下降的表观矛盾。研究证实,外排泵协同激活是微生物响应OTC胁迫的重要策略,且通过运行参数优化(如曝气压调控)可使TN去除率恢复至87.4±5.8%。综上,MABR系统在处理含抗生素废水时展现出高效脱氮能力与显著过程韧性。
致谢:本研究得到国家自然科学基金基础科学中心项目(52388101)、广东省引进创新创业团队计划和广东省杰出青年科学基金的资助。


 粤ICP备05008833号
粤ICP备05008833号
